
輪形動物としても知られるワムシは、輪形動物門に属する微小な水生動物です。その名前は頭部に繊毛冠を持つことに由来し、移動と食物の捕集に使う特徴的な構造をしています。
繊毛冠は脈動する繊毛でできていて、回転する車輪のように見えることからこのように呼ばれています。
2,000種を超えるワムシは、世界中で見つけられます。主な生息地は池や湖などの淡水です。また、藻類、地衣類、土壌、下水、海洋環境、永久凍土層、さらには他の動物の体内外で見つかることもあります。

顕微鏡観察のためのサンプルとなるワムシの採取。画像提供:Chloé Savard氏。
寄生性のあるワムシ種は少数ありますが、ほとんどのワムシは自由生活で、プランクトン間を泳ぐもの、堆積物内をはい回るもの、水中の水生植物に付着するもの、分泌されたゼラチン状の棲管内部に住むものがいます。これらの小さな無脊椎動物は、さまざまな環境圧(捕食など)に適応するために多様な形態と移動方法を発達させた、極めて多様な動物群を形成しています。
ワムシの一般的な生態
ワムシは単細胞生物でしょうか、それとも多細胞生物なのでしょうか。一見、ワムシは単細胞微生物のように見えます。多くの単細胞原生生物より小さいにもかかわらず、実際には約1,000の細胞から形成されています。
ワムシの体長は最大2mmになることがありますが、通常は0.1~0.5mmです。存在する最小のワムシは0.006~0.008mm(6~8µm)ほどで、これは人間の赤血球の大きさの6倍にすぎません。

微分干渉観察(DIC)で撮影されたハオリワムシ属の一種画像提供:Chloé Savard氏。
ワムシの形態にはさまざまなものがありますが、主な体のつくりは同じです:頭(繊毛冠)、体(胴部)、足からなります。足(存在する場合)には足指を持つ場合があります(通常2本)。最大4本の足指を持つ種もあれば、まったく持たない種もあります。
種によっては、足が自由に伸縮するものもあります。この現象は特にヒルガタワムシに見られますが、Rotaria neptuna種にも見られます。粘着性物質を分泌する足腺を足に持つ場合もあります。ワムシはこの物質を接着剤として使い、植物、岩、木片、動物などさまざまな基質に一時的に付着します。
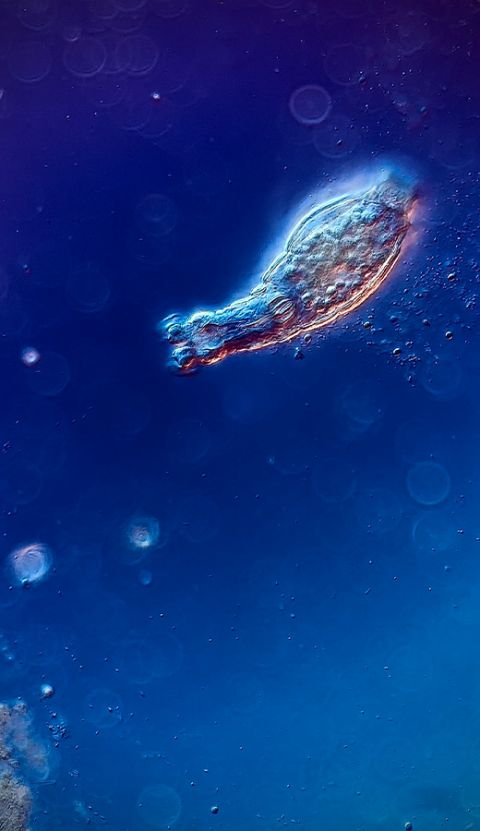
DICを使い200倍で撮影されたヒルガタワムシ。画像提供:Chloé Savard氏。
浮遊種と自由遊泳種の多くは足がありません。固着性のワムシもいます。幼生期に泳ぎ回って完璧な基質を見つけると、そこに付着して一生過ごします。この例がStephanoceros fimbriatusです。
これらの原始的生物には循環器系はありませんが、脳、小さな赤い目、完全な消化管、筋肉系、生殖器系、排泄系を持ちます。
胴体内にあるほとんどの臓器系を保護するため、ワムシが持つ表皮とも呼ばれる体壁は、非常に厚く硬いものも薄く柔らかいものもあります。硬い殻を持つ種は被甲のあるワムシ、柔らかい種は被甲のないワムシと呼ばれます。
被甲のあるワムシの中には、表皮の前端または後端にとげを持ち、捕食者への防御手段とする種もあります。これはKeratellaやAsplanchnaの種に見られ、カイアシなどの無脊椎捕食者が発する可溶性化学信号に反応してとげが生成されます。Keratellaの生息環境に捕食者が多く存在するほど、化学信号も多くなり、これらのワムシのとげが発達する傾向があります。
ワムシの頭部には2つの繊毛環を持つ繊毛冠があり、そこから作られる水の渦を使って食物を集めたり水柱内を泳ぎ回ったりします。
緩歩動物のように、ヒルガタワムシはクリプトビオシスと呼ばれる休眠状態になることができます。より具体的には乾燥無代謝休眠です。この状態になると、長期間にわたり体内から水分が完全に失われる乾燥を生き延びることができます。
ヒルガタワムシは乾燥無代謝休眠の状態になって、菌の寄生を逃れたり、極寒や電離放射線への暴露を耐え抜いたりできます。豆知識:2021年7月に発表された研究によると、北極の永久凍土から採取されたヒルガタワムシが、2万4千年にわたる凍結を経て乾燥無代謝休眠から復活しました。
ワムシは何を食べるか
極小甲殻類と原生生物とともに、ワムシは淡水動物プランクトンを支配する第3群を形成します。有機堆積物、細菌、酵母、藻類、小さな繊毛虫類、その他の原生生物からなる食事に加えて、環境状態と捕食量に応じて採食行動を適応させることができます。
ワムシの中には、小さなワムシを捕食するほど大きな種もあります。

DICを使い100倍で撮影されたStephanoceros fimbriatus。 画像提供:Chloé Savard氏。
ワムシは繊毛冠を使って水の渦を作り出し、食物をとらえると、口に直接取り込みます。口に入った食物は、そしゃく嚢と呼ばれる咽頭が変化した器官で押しつぶされます。この器官はワムシ独特のものです。食物をかみ砕くとき、そしゃく嚢は脈打つ心臓のように見えるため、循環器系を一切持たないワムシを一見すると勘違いする場合があります。
筋肉でできたそしゃく嚢には、口器と呼ばれるあごのような構造があり、さまざまな食物をとらえ、つかみ、突き刺す役割を果たします。口器の構成は種によって異なるため、ワムシを特定する際に役立ちます。押しつぶされた食物は口器を出ると、食道を下って胃、腸、最後に肛門に到達し、体から出ます。
ワムシ自体も、原生生物(特に繊毛虫類)、昆虫、他のワムシ、枝角類、カイアシ、魚など、たくさんの水中捕食者に捕食されます。ワムシは捕食者から逃れるため、ジャンプする動き、粘液鞘、とげ、厚い殻など、さまざまな方法を発展させました。
ワムシの生殖方法
ワムシの生殖方法は輪形動物門内で大いに異なります。ワムシは有性生殖も無性生殖も行います。どちらの場合も卵を抱く必要があります。
有性生殖を行う種では、雄と雌の両方が個体群内に見られます。雌のワムシが産む両性生殖卵は単数体であり、雄の精子で受精させる必要があります。卵が受精しない場合、単為生殖で雄に成長します。有性生殖を行うプランクトン種では性的二形がよく見られ、足を持たない雄のワムシは雌のワムシよりも小さく、速く泳ぎます。
一方、ヒルガタワムシは単為生殖により無性生殖します。個体群を構成するのは雌のみです。さらに、雌の幼生ワムシが産まれるように雄の精子で受精させる必要はありません。このような種類の卵は倍数性単為生殖卵であり、2倍体です。
ヒルガタワムシは、枝角類、アブラムシ、ハチ、アリなどの種とともに、雄のいない数少ない動物群の1つです。ヒルガタワムシはおよそ4千年もの間、雌だけの動物群として発達してきたのです。
ワムシを顕微鏡で観察する方法
ワムシはほぼどこでも見つけられます。池、湖、あるいは建物の側溝からでもサンプルを採取すると、少なくとも1種のワムシが顕微鏡で見つかります。

顕微鏡観察のためのサンプルとなるワムシの採取。画像提供:Chloé Savard氏。
ヒルガタワムシは特に藻類や地衣類に豊富に存在します。休眠状態の可能性が高いので、藻類や地衣類を24時間水に浸してから絞り、ペトリ皿に入れて顕微鏡で観察します。
ヒルガタワムシは泳ぐよりはい回っていることが多いため、初めて見る人は虫と勘違いするかもしれません(私が初めて見たときが正にそうでした)。ワムシを最適に観察するには、複式顕微鏡で明視野または暗視野照明を使用します。
参考文献
Allen, A. A. (1968). Morphology of the planktonic rotifer Polyarthra vulgaris. Transactions of the American Microscopical Society, 60-69.
Bogdan, K. G., & Gilbert, J. J. (1982). Seasonal patterns of feeding by natural populations of Keratella, Polyarthra, and Bosmina: Clearance rates, selectivities, and contributions to community grazing 1. Limnology and Oceanography, 27(5), 918-934.
De Paggi, S. B. J., Wallace, R., Fontaneto, D., & Marinone, M. C. (2020). Phylum Rotifera. Thorp and Covich’s Freshwater Invertebrates, 145–200.
Gladyshev, E., & Meselson, M. (2008). Extreme resistance of bdelloid rotifers to ionizing radiation. Proceedings of the National Academy of Sciences, 105(13), 5139-5144.
Gribble, K. E., & Snell, T. W. (2018). Rotifers as a Model for the Biology of Aging. Conn’s Handbook of Models for Human Aging, 483–495.
Hickman, C. P., Roberts, L. S., Larson, A., Ober, W. C., & Garrison, C. (2015). Animal
diversity. WC Brown. 183.
Ricci, C. (1998). Anhydrobiotic capabilities of bdelloid rotifers. Hydrobiologia, 387, 321-326.
Segers, H., & De Smet, W. H. (2007). Diversity and endemism in Rotifera: a review, and Keratella Bory de St Vincent. Protist Diversity and Geographical Distribution, 69-82.
Shmakova, L., Malavin, S., Iakovenko, N., Vishnivetskaya, T., Shain, D., Plewka, M., & Rivkina, E. (2021). A living bdelloid rotifer from 24,000-year-old Arctic permafrost. Current Biology, 31(11), R712-R713.
Taylor, R. A. J. (2019). Other invertebrates. Taylor’s Power Law, 305–326.
Wallace, R. L. (2002). Rotifers: exquisite metazoans. Integrative and Comparative Biology, 42(3), 660-667.
Wallace, R. L., Snell, T. W., Walsh, E. J., Sarma, S. S. S., & Segers, H. (2019). Phylum Rotifera. Thorp and Covich’s Freshwater Invertebrates, 219–267.
Welch, D. B. M., & Meselson, M. (2000). Evidence for the evolution of bdelloid rotifers without sexual reproduction or genetic exchange. Science, 288(5469), 1211-1215.
Wininger, J. D. (2004). Parthenogenetic Stem Cells. Handbook of Stem Cells, 635–637.
