
Die Fluoreszenzbildgebung war und ist seit jeher ein wichtiges Hilfsmittel in der biologischen Forschung. Sie ermöglichte es unzählige Male, zelluläre Strukturen und dynamische Prozesse auf mikroskopischer Ebene sichtbar zu machen.
Jeder Anwender eines Fluoreszenzmikroskops kennt das schwache Hintergrundsignal, das bei der Abbildung biologischer Proben überall im Gewebe zu sehen ist. Die Intensität des Signals kann bei verschiedenen Strukturen variieren und sich je nach Wellenlänge der Anregungsquelle verändern.
Bevor jedoch Protokolle und Aufzeichnungen auf Fehler in der Färbetechnik überprüft werden, empfiehlt es sich, zunächst einmal die biologische Zusammensetzung der Probe zu überprüfen. Ist sie zellularisiert? Pigmentiert? Sind viele Strukturproteine vorhanden? Wenn eines davon der Fall ist, ist das Signal möglicherweise nicht so stark durch äußere Faktoren bedingt wie angenommen. Vielmehr kann es sich bei diesem Hintergrundsignal ganz oder teilweise um natürliche Autofluoreszenz handeln.
Das innere Leuchten in der Biologie – was ist Autofluoreszenz?
Autofluoreszenz ist die natürliche Emission von Licht durch biologische Strukturen und ein weit verbreitetes Phänomen in Zellen und Gewebe. Sie wird durch endogene molekulare Komponenten verursacht, die fluoreszieren können. Ähnlich wie technische Fluorophore, die für Färbungen und Farbstoffe verwendet werden, bestehen autofluoreszierende Moleküle meistens aus polyzyklischen Kohlenwasserstoffen mit delokalisierten Elektronen, die durch einfallende Photonen angeregt werden können. Autofluoreszierende Moleküle sind resistent gegen eine effiziente Schwingungsrelaxation, wenn sie durch einfallendes Licht angeregt werden. Infolgedessen wird die überschüssige Energie als neues Photon emittiert, das über eine geringere Energie und eine höhere Wellenlänge als das anregende Photon verfügt. Auch wenn die Mechanismen vertraut klingen, stellt sich vielleicht die Frage, warum man Autofluoreszenz nicht überall im Leben begegnet. Schließlich ist der Mensch selbst eine biologische Einheit. Dennoch lässt sich unsere Fluoreszenz nicht direkt erkennen.
Einige endogene Moleküle (wie Chitin) sind dafür bekannt, dass sie unter ultravioletter Anregung autofluoreszieren (siehe Abbildung 1, links). Im Gegensatz zu ihren künstlich hergestellten Pendants im Labor werden viele Autofluorophore jedoch nicht so einfach durch einfallende Photonen angeregt. Zudem mag zwar in einem Färbeprotokoll die Konzentration von künstlichen Kontrastmitteln angepasst werden können, um ein Fluoreszenzsignal zu dämpfen oder zu verstärken, aber die natürliche Autofluoreszenz ist immer auf biologische Konzentrationen beschränkt.
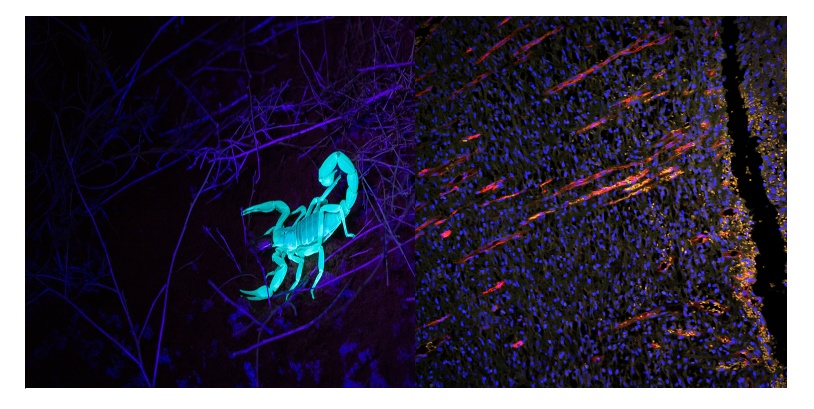
Abbildung 1: Beispiele für Autofluoreszenz in lebenden Proben. Links: Autofluoreszenz eines Skorpions bei Anregung mit UV-Licht. Rechts: Neovaskulatur in der Haut einer Ratte mit vertikalem Wachstum in Richtung des Wundschnitts in der epidermalen Schicht im Rahmen des Heilungsgeschehens 10 Tage nach Herbeiführung des Traumas. Blau (DAPI): Zellkern. Orange (Autofluoreszenz): Hautgewebe. Rot (CD31): Blutgefäße. Bildaufnahme mit dem Weitfeld-Fluoreszenzmikroskop BX51 von Olympus und der DP71 Kamera. Bildquelle: LiShuang Li, Experimental Research Center, China Academy of Chinese Medical Sciences.
Autofluoreszierende Moleküle haben außerdem distinkte Anregungsfenster, wenngleich die meisten Autofluoreszenzen im UV-grünen Bereich angeregt werden und relativ breite Anregungsspektren ergeben. All diese Faktoren zusammen, führen dazu, dass Autofluoreszenz viel häufiger auftritt, wenn Bildgebungssysteme mit speziellen fluoreszenzanregenden Lichtquellen und Photonendetektoren verwendet werden, die empfindlicher sind als das bloße Auge (Abbildung 1, rechts: orangefarbenes Signal).
Häufige Quellen von Autofluoreszenz in der biowissenschaftlichen Forschung
Eine Erklärung des Vorhandenseins von Autofluoreszenz bei Bildgebungsexperimenten in der Forschung liefern bestimmte biologische Komponenten der Probe. Hier sind einige Quellen von Autofluoreszenz, die bei der wissenschaftlichen Arbeit häufig eine Rolle spielen:
1. Nikotinamidadenindinukleotid
Nikotinamidadenindinukleotid oder NAD(P)H ist ein metabolischer Cofaktor und Elektronenträger, der im gesamten Zytoplasma vorkommt und an der Glykolyse und am Pentosephosphatweg beteiligt ist. Da das Molekül eine wichtige Komponente des Zellstoffwechsels ist, ist NAD(P)H-bedingte Autofluoreszenz in fast allen lebenden Zellen vorhanden. Um seine Stoffwechselfunktionen zu erfüllen, existiert das Molekül in einem oxidierten (NAD+) und in einem reduzierten (NAD(P)H) Zustand. Allerdings ist nur NAD(P)H fluoreszenzerzeugend. Das oxidierte Gegenstück NAD+ weist keine Autofluoreszenz auf. Anregung: 340 nm; Emission: 450 nm. (Quelle:Chance et al. 1979; Georgakoudi et al. 2002).
2. Flavin
Flavinadenindinukleotid (FAD) ist ein metabolischer Cofaktor, der eine wichtige Rolle im TCA-Zyklus und in der Elektronentransportkette spielt. Er ist hauptsächlich in den Mitochondrien lokalisiert, wo aktive Stoffwechselprozesse Hotspots von Autofluoreszenzsignalen erzeugen. Einige Wissenschaftler bezeichnen die Autofluoreszenz von Flavin als „Flavoprotein-Fluoreszenz“, da FAD funktionell an Proteinkomplexe in den Mitochondrien (wie Succinatdehydrogenase) gebunden ist. Anders als bei NAD(P)H fluoresziert nur die oxidierte Form von FAD, während die reduzierte Form nicht fluoreszierend ist. Anregung: 380–490 nm; Emission: 520–560 nm. (Quelle:Chance et al. 1979; Deylet al. 1980).
3. Kollagen
Kollagen ist ein wichtiges mechanisches Protein mit Stützfunktion in den meisten Geweben. Es kommt in der Dermis der Haut, der extrazellulären Matrix innerer Organe und dem umliegenden Gefäßsystem vor. Außerdem ist es auch in Sehnen, Bändern, Haaren und Fingernägeln enthalten. In Zellkulturen ist Kollagen nur selten zu sehen, aber bei der Darstellung von In-vivo-Proben oder ganzen Geweben ist Kollagen unvermeidlich. Anregung: 270 nm; Emission: 390 nm. (Quelle:Georgakoudi et al. 2002).
4. Elastin
Ein weiteres wichtiges Protein in der extrazellulären Matrix (ECM) ist Elastin, das häufig mit Kollagen durchsetzt ist, um der ECM stärkere mechanische Dehnbarkeit zu verleihen. Elastin ist dicht an Gefäße angelagert, die als Anpassung auf Blutdruckveränderungen einer häufigen elastischen Verformung unterliegen. Es ist auch in der Haut zu finden, die elastisch sein muss, um sich der Bewegung der darunter liegenden Knochen und Muskeln anzupassen. Die Autofluoreszenz von Strukturproteinen wie Elastin spielt wiederum bei der Darstellung von In-vivo-Proben oder Gewebeproben eine Rolle. Anregung: 350–450 nm; Emission: 420–520 nm. (Quelle:Deylet al. 1980).
5. Lipofuszin
Lipofuszin ist ein autofluoreszierendes Molekül, das einen kleinen, aber nicht unrelevanten Teil der Fluoreszenzspektren von Neuronen, Gliazellen, Skelettmuskelzellen und Herzmuskelzellen ausmacht. Es ist in Zellkulturen und auch in ganzen Geweben zu erkennen. Offensichtlich kommt es mit zunehmender biologischer Alterung einer Probe immer deutlicher zum Vorschein. Obwohl der Name vermuten ließe, dass ein Lipid oder Lipoprotein dafür verantwortlich ist, wird das Autofluoreszenzsignal von Lipofuszin auf ein Gemisch aus Proteinen, Kohlenhydraten und Lipiden zurückgeführt. Anregung: 345–490 nm; Emission: 460–670 nm. (Quelle:Billinton und Knight 2001).
6. Tryptophan
Tryptophan ist als essenzielle Aminosäure ein Baustein für Signalmoleküle (wie Serotonin und Melatonin), die in der neurologischen Forschung eine Rolle spielen, und ein wichtiger Bestandteil der Proteinbiosynthese. Es ist quasi omnipräsent, da Tryptophanreste in den meisten aufgefalteten Proteinen zu finden sind, daher durchdringt sein Autofluoreszenzsignal Zellen und Gewebe gleichermaßen. Da die Strukturchemie von Tryptophan an die Proteinkonformation gebunden ist, verändern sich Wellenlänge und Intensität der Autofluoreszenz in Abhängigkeit von Veränderungen der Proteinstruktur und des Bindungsstatus. Anregung: 280 nm; Emission: 350 nm. (Quelle:Ghisaidoobe und Chung 2014).
7. Melanin
Melanin ist ein natürliches Pigment, das zur Haut-, Haar- und Augenfarbe beiträgt. Melanin wird von Zellen in der Basalschicht der Epidermis der Haut produziert und wirkt als Lichtschutzmolekül, das verhindert, dass UV-Licht die wertvollen Proteine und die DNA in den nach außen zeigenden Hautzellen schädigt. Die natürliche Konzentration und Verteilung von Melanin können sogar innerhalb ein und derselben Probe variabel sein, was bei der direkten Kultur von Melanozyten unerheblich ist, aber bei Experimenten mit Blick durch die Haut hindurch berücksichtigt werden muss. Anregung: 340–400 nm; Emission: 360–560 nm. (Quelle:Gallas und Eisner 1987).
Fluoreszenz aufgrund der Probenvorbereitung
Die oben genannten Autofluoreszenzquellen emittieren Photonen auf natürliche Weise aus biologischem Gewebe. Oftmals kommt es auch zu Fluoreszenz, die von nichtbiologischen Bestandteilen oder Reagenzien stammt, die zur Vorbereitung von Proben im Labor benötigt werden.
So können beispielsweise Kunststoffböden von Petrischalen, Mikrotiterplatten und Zellkulturflaschen in einem breiten Spektrum hell fluoreszieren. Zur Fluoreszenzbildgebung von biologischen Proben empfehlen wir daher die Verwendung von Behältern mit Glasboden oder speziellem nicht fluoreszierendem Polymer. Auch Phenolrot, ein gängiger Zusatzstoff in Zellkulturmedien, kann die Hintergrundfluoreszenz bei der Abbildung von Lebendzellen deutlich erhöhen. Dieser Effekt lässt sich leicht verhindern, indem man vor Beginn der Bildgebungsexperimente das Medium durch eine phenolrotfreie Alternative ersetzt.
Papieretiketten oder Aufkleber können ähnliche Probleme verursachen, da Papier stark fluoreszierend ist. Wenn Aufkleber auf Behältern oder Objektträgern verwendet werden, sollten diese bei der Abbildung der Proben ausgelassen werden.
Auch Aldehyd-Fixiermittel wird häufig in Färbeprotokollen verwendet. Reagenzien wie Glutaraldehyd oder Formaldehyd reagieren mit Proteinen und erzeugen fluoreszierende Vernetzungen in Zellen und im Gewebe. Zur Vermeidung einer Akkumulierung unerwünschter Fluoreszenzsignale empfiehlt es sich, diese Reagenzien durch aldehydfreie Fixiermittel zu ersetzen.
Auch wenn es sich bei diesen Beispielen nicht um natürlich vorkommende Autofluoreszenzquellen handelt, können durch die Kenntnis ihrer Herkunft und ihrer Auswirkungen unerwünschte Einflüsse auf Bilddaten verhindert werden.
Umgang mit unerwünschter Autofluoreszenz in einem Bildgebungsexperiment
Vorangehend wurden einige Autofluorophore und ihre Quellen beschrieben. Nun stellt sich folgende Frage: Was kann gegen diese Signale getan werden, bevor ein Bildgebungsexperiment begonnen wird?
Bei Gewebe mit einfacher Struktur oder monozellulären Kulturen sind die Anregungs- und Emissionsspektren handelsüblicher Färbemittel bedacht auszuwählen und die entsprechenden schmalen Filter so zu wählen, dass größere Autofluoreszenzspitzen vermieden werden. So kann das Signal-Rausch-Verhältnis erheblich verbessert werden.
Wenn das Protokoll es zulässt, sollten Färbemittel mit hoher Quanteneffizienz gewählt oder eventuell höhere Konzentrationen des Kontrastmittels ausprobiert werden. Dadurch kann sich auch das Signal verstärken, allerdings zu höheren Kosten. Für moderne Strategien wird Anregungslicht im Nahinfrarotbereich (NIR) mit Markern verwendet, die über 700 nm anregbar sind (z. B. Cy7 oder Alexa Fluor 750), um die Anregungs- und Emissionsbereiche häufig vorkommender Autofluorophore zu vermeiden (Abbildung 2).
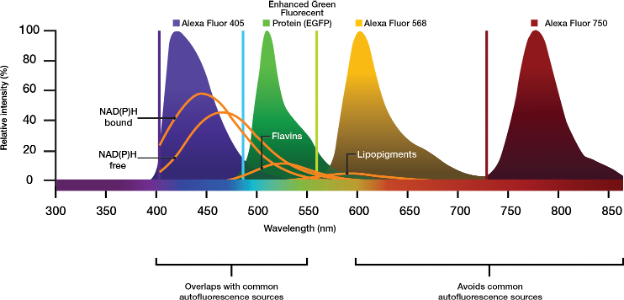
Abbildung 2: Emissionspeaks von kommerziellen Alexa-Fluorophoren relativ zu autofluoreszierenden Molekülen, die in der biologischen Bildgebung üblich sind. Mit der Auswahl von Kontrastmitteln, die im NIR-Bereich anregen und emittieren, können unerwünschte Emissionen von Autofluoreszenzquellen vermieden werden.
Eventuell könnten auch alternative Bildgebungsmodalitäten für Experimente infrage kommen. Bei dicken Proben oder dickem Gewebe können konfokale oder Multiphotonen-Mikroskopsysteme den Gesamtanteil der Autofluoreszenz reduzieren. Sie tun dies, indem sie die Ansammlung von unscharfem Licht unterbinden bzw. die Anregung auf die Brennebene beschränken.
Mit der Biolumineszenz-Bildgebungstechnik kann beispielsweise ausgeschlossen werden, dass ein von Autofluorophoren stammendes Signal in die erfassten Daten aufgenommen wird. Lumineszenzexperimente erfordern kein Anregungslicht für die Chemilumineszenzreaktionen, um Emissionsphotonen zu erzeugen, sodass Autofluorophore gar nicht erst angeregt werden.
Wenn der Anteil des Autofluoreszenzsignals vor der Bildgebung nicht verringert werden kann, können Bildverarbeitungstechniken nach der Aufnahme (wie spektrale Entmischung oder Hintergrundsubtraktion) effektive Alternativen bieten. Diese Computertechniken erfordern, dass Wissenschaftler die endogenen Fluorophore in den Proben herausfiltern oder die Autofluoreszenzspektren kennen, um diesen Anteil wirksam zu entfernen.
Ähnliche Artikel
Bildgebende Verfahren zur Beurteilung der Wirksamkeit von Arzneimitteln
